| |
This article seeks to provide professionals and lay people with a meaningful understanding of how and why human beings experience a variety of anomalous states of feeling, knowing, healing, and being that are usually associated with paranormal, mystical, religious, spiritual, transcendental, and biogenetic evolutionary states of felt experience and intuitive consciousness. I propose to clarify these biogenetic and anomalous experiential states by introducing the following: 1) the basic structure of the human nervous system, 2) the nature and function of neuron cell potential, 3) the constant spread of the biogenetic DNA potential, and 4) the hidden variable of quantum transmission potential. This paper also seeks to address interactive relational qualities that seem to exist between psychological, neurological, molecular, and quantal processes. These interactive biogenetic and psychoneuroquantum potentials were developed in a manner consistent with what we know thus far in the fields of depth psychology, neurophysiology, DNA molecular biology, and quantum unified field physics. It is my contention that an understanding of these biogenetic and psychoneuroquantum potentials is necessary to the progress of a more complete science of the human nervous system. I believe that such a radically extended nervous system model is not only necessary as a preliminary explanation and clarification of the vast depth of experience within ourselves, but also as a visionary direction for future scientific research that can impart new meaning to processes operative deep within the nervous system that cannot be directly observed by our senses or by the present technological extension of our senses. I believe that the progress of our knowledge regarding human nervous system functioning rests on the crucial interplay between rational analysis and intuitive cognition. The faculty of intuitive cognition alone can leap beyond the analysis of empirical data to show us where to look for new facts, but it does not have the logical precision to connect the potential facts with the given data of empirical research. The faculty of reason alone, on the other hand, has the logical precision to integrate intuitive potential data with empirical research data, but it cannot leap far beyond the analysis of the given data of empirical research to show us the radical future paradigms of humanity and reality. Therefore, I hold that without the complementary interplay between reason and intuition a science of the human nervous system cannot be a truly progressive science, for the progress of any science is both a step by step analysis of present data and an intuitive visionary leap into our potential understanding and realization of "humanity's psychoneuroquantum potential."
In order to communicate the psychoneuroquantum potential of human nervous system functioning, I use a multi-disciplinary approach that employs terminology and general references from biogenetics, quantum physics, neurology, and depth psychology. Using this multi-disciplinary approach, I develop an integrated nervous system model which gives meaning and validity to experiences that are as yet unquantifiable and unapproachable within the current nervous system paradigm in our medical and behavioral sciences. In this way, I hope to promote a more holistic view and understanding of how we, as human beings, can experience a psychoneuroquantum potential that has the capacity to promote health, healing, meaning, purpose, and growth amidst the mundane activities and relationships of our daily lives. By remaining open to this human psychoneuroquantum potential, we may begin to scientifically approach and validate not only mystical, religious, and paranormal experiences, but also the daily human experiences and behaviors associated with embodied faith, love, wonder, empathy, meaning, purpose, compassion, generosity, gratefulness, peace, wisdom, awe, and kindness.
The Psychoneuroquantum Body of Original Faith
Central to all the major religions of the world is the ultimately valued experience of embodied Faith. It is my hypothesis that most of the established world religions have misconstrued faith to mean the unquestioning acceptance of, identification with, and attachment to a particular historical figure, creed, dogma, text, symbol, institution, or belief. These world religions do not readily acknowledge that, though their beliefs and symbol systems are varied and unique, all human beings actually experience the same inner quality. integrity, and meaning of embodied faith regardless of their religious affiliation.
The innate experience of embodied faith, in every religion, is a common motivating factor in the behavioral practice and expression of love, compassion, wisdom, truth, generosity, respect, appreciation, sincerity, humility, and empathy. Based on the premise that the same experiential quality and meaning of embodied faith is common to all human beings, one cannot logically ask people from another religion, "What is your faith?" Because the experience of Faith is "Only One", there cannot be another to compare it to. It would be more accurate to ask a person, "What are your religious beliefs and symbols that arouses your felt embodiment of faith and how does this felt experience of faith impact the practice of compassion and wisdom in your life and relationships?" This would help distinguish the variety of beliefs and symbol systems from the commonly shared experience of embodied Faith and its function as a universal motivating factor in interpersonal behaviors and experiences that are valued worldwide.
As a meditation psychologist, it is my lived hypothesis that this innately felt experience of Faith, common to all human beings from differing religious systems, is fundamentally dependent on the psychoneurological emergence of an innate quantum informational potential operative deep within the human nervous system. Within this embodied psychoneuroquantum perspective, I define Faith as the felt experience of a deep and abiding confidence in a Vast Wisdom that penetrates our bodily self, the activities and relationships of our life, and the Universe as a Whole. This natural human feeling of embodied Faith fills one with a deep sense of Unity, Order, Meaning, Purpose, Wonder, Compassion, and Mystery. I posit that this Original Faith, or deep confidence in a Vast Wisdom penetrating the Body and All Existence, is actually an Innate Psychoneuroquantum Potential that, when realized and integrated through the felt experience of the body, establishes a confident biological awareness of our human need completion, spiritual life fulfillment, and total death entrustment.
In summing up what I have said thus far, the bodily emergence and continuation of Original Faith, as a felt psychoneuroquantum experience, is not dependent on any belief, idea, or symbol system. This is precisely so because the felt bodily meaning of Original Faith arises from a psychoneuroquantum depth of human neurological functioning. Original Faith is not like things that we can acquire, own, or gain. It is innate and arises as our immediate felt experience before any language, culture, race, history, religious views, knowledge, and thinking mind. In other words, the Precious Source and infinitely profound life meaning of Original Faith is preverbal, prereflective, precultural, prereligious, and prehistorical
When the experience of Original Faith is viewed within the context of our psychoneuroquantum potential, we can see that it cannot be created or owned by any specific religious or spiritual symbol or belief. Further, the felt emergence of this psychoneuroquantum potential and its integration into conscious bodily awareness fulfills the basic human need for a sense of deep confidence in the Vast Wisdom that penetrates both life and death while providing humanity with the capacity to see through the illusory nature of their individual personalities, egos, cultures, religions, languages, and racial histories. Thus, upon the fulfillment of this psychoneuroquantum potential rests the hope and the future of ONE Humanity----a Humanity that encourages and celebrates unique differences amongst all human beings while discouraging personal, political , religious, and economic beliefs, views, and behavioral practices that foster hate, arrogance, greed, ignorance, superiority, exclusiveness, and intolerance.
General Orientation to the Human Nervous System
The evolutionary emergence of the human nervous system manifests the organizational complexity of the life process by the ordering of some twenty six billion electrochemical components in a distinctly coherent and purposive manner. This most complex biological system constitutes a multidimensional tool specifically designed to help us function in the environment we find ourselves in. It is the ultimate instrument of adaptation which enables us "to do something.. in the most pragmatic sense of that phrase. The nervous system has helped us to adapt the external environment to our constitutional needs and this accounts for our evolutionary progress as a species.
The human nervous system functions as a whole, but for descriptive purposes it can be divided into two categories, the central nervous system and the peripheral nervous system. The central nervous system is composed of the brain and spinal cord. These nervous organs are protected by the bony case of the skull and the spinal column, respectively. The most distinctive feature of the central nervous system and the most conspicuous evolutionary difference is its magnificent brain. A human being has more innervation potential within the cranial cavity than any other of the higher mammals. Indeed, the human brain has about one-half of the nerve cells in the entire nervous system. It has so expanded into its allotted cranial space that is shows many ridges and foldings of the surface. This greater centralization of nerve cells in the brain provides a human being with an enormous potential for sophisticated calculation that in turn permits the accomplishment of complex manipulations of the environment. (Klemm, 1972)
One of the main functions of the spinal cord is to conduct nervous signals to and from the brain. Although sensory messages from the head region enter and leave the brain directly via the cranial nerves, sense data from the rest of the body is conducted to the brain via the spinal cord. In the same respect, the control of the skeletal-musculature, aside from the head and neck region, is exercised by the brain through the nervous pathways running within and coursing outside of the spinal cord. The spinal cord also serves as a reflex center which operates according to a closed-circuit response. This kind of reflex circuitry makes possible an array of automatic and instantaneous responses to certain sensory signals. Such reflex circuitry can initiate a localized motor response in the segment of the body that received the sensory stimulation without cerebral participation. (Guyton, 1972)
Diagram of Central Nervous System
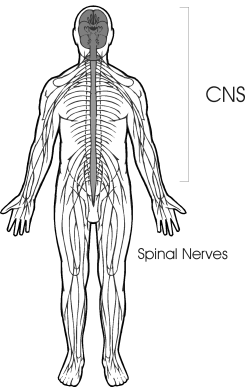
The network of nervous pathways leaving the brain and spinal cord constitute the peripheral nervous system. This peripheral system is composed of chains of nerve cells that course like roots throughout the biological soil. The network of root nerves leaving the brain provide the information concerning the head and neck regions, while the network of root nerves leaving the spinal cord provide the necessary information concerning the state of the internal organs and the surface regions of the body. This peripheral network is subdivided into the somatic and autonomic nervous systems. The autonomic system consists of nerve cells which innervate the internal organs, the involuntary musculature, and the secretions of many glands.
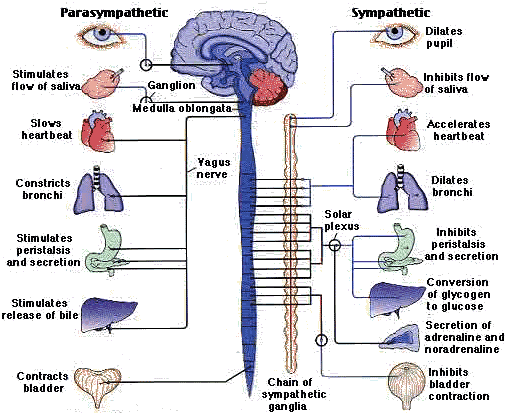
The autonomic system itself is further divided into the sympathetic system and the parasympathetic system. These two autonomic systems generally exert opposing influence on the same organs, glands and muscles. The sympathetic system, for example, is responsible for contraction of arteries, acceleration of heart beat, inhibition of digestion, and dilation of the pupils. The parasympathetic system, on the other hand, is responsible for the expansion of arteries, deceleration of heart beat, activating secretions in the stomach, and constriction of the pupils. The opposing biological effects of these autonomic systems have become the basis for the usually accepted generalization that the sympathetic system mobilizes and activates body resources for immediate energy expenditures, while the parasympathetic system tends to relax and conserve body resources for future energy expenditures. Although the biological processes associated with the sympathetic and parasympathetic systems are usually beyond voluntary control, they need not be necessarily so. This is to say that voluntary control is latent or potential in the autonomic nervous system. (Ferguson, 1973; Schwartz, 1973)
The somatic nervous system is responsible for sensory reception and bodily action. This system is composed of afferent sensory pathways and efferent motor pathways. The former afferent neural pathways are made up of linked nerve cells that transmit excitations received from the sensory receptors deep within the anatomy and at the body periphery to the central nervous system. The latter efferent neural pathways are made up of linked nerve cells that transmit excitations from the central nervous system to the glands and muscles. The nervous receptor mechanisms that collect sensory information can be classified into three different physiological types of specialized cells or modified nerve fibers: 1) exteroceptive receptors are usually associated with the five peripheral senses of sight, touch, taste, smell and hearing; 2) interoceptive receptors are sensitive to the internal states of the anatomy, including the deep sensations of viscera, tendons, muscles, joints, and bones; 3) proprio-ceptive receptors located in the inner ear and skeletal muscles collect information regarding the body's position and equilibrium in space. The various sensory receptors are primarily responsible for the transformation of physical energies into nervous impulses. Once this transformation takes place, the impulse signals can travel over the afferent pathways of the peripheral nervous system to the spinal reflex centers or to the higher computational areas in the brain. Finally, impulse signals are sent from the central nervous system across the efferent pathways of the peripheral nervous system to the appropriate muscles and/or glands. This, of course, is an oversimplification of what happens during sensory Conduction and motor transmission, for there is a constant interaction between all Parts of the nervous system. (Guyton, 1972; Noback, & Demarest, 1975)
Nature and Function of Neuron Cell Potential
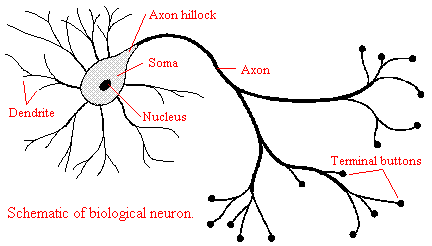
A nerve Cell may be divided into three major physiological parts, the soma, which is the main cell body; the axon, which is the fairly thick and lengthy fiber that extends from the soma;, and the dendrites, which are relatively short, root-like projections of the main cell body. The dendrites serve to receive messages from receptors or other nerve cells, whereas the axon transmits messages to other nerve cells or to the effector cells in muscles and glands. The soma is in charge of manufacturing the biochemical substances that keep the whole Cell alive while also serving as a conduction path between the dendrites and the axon.
The multitude of nerve cells which comprise the nervous system may be divided into three functional categories: 1` sensory neurons that are responsible for detecting changes in certain features of the inner and outer environment; 2)interneurons which are primarily responsible for processing data transmissions by exciting each other in a sequential fashion; and 3) motor neurons that are directly responsible for a specific behavioral response by making the glands secrete and the muscles contract. All three categories of neurons or nerve Cells exhibit a variety of biochemical processes when performing their peculiar functions and when they are at rest. Many of these processes occur in all the other kinds of Cells that make up the body. Like other living cells, neurons evidence the fundamental processes of cellular metabolism ?? the chemical changes by which energy is provided for the vital biological activities associated with the build-up and destruction of protoplasm. However, the neuron is a specialized cell that manifests a number of unique electrochemical activities that are not found in the other cells of the body.( Schwartz, 1973; Thompson, 1975)
The unique characteristic of neurons is clearly associated with their semipermeable membrane that is in immediate contact with an external biochemical fluid. This fluid substance contains electrically charged molecules Called ions. The porous nature of the neuron membrane allows negative ions to pass in and out of the cell freely while restricting the passage of most positive ions. As a result, there are more positive ions than negative ions outside the cell membrane and more negative than positive ions inside of the Cell membrane. This is as much to say that a positive electrical charge predominates in the extracellular environment while a negative electrical charge predominates in the intracellular environment. Such an electrical polarization creates a measurable electrical potential, the resting potential, across the neuron membrane. This resting potential is labeled as a negative neuron potential because the cell itself is more negative than the surrounding biochemical fluid. The negative potential across the cellular membrane is typically about minus seventy millivolts that is one fifth of the potential difference of a common flashlight battery. (Mishlove, 1975; Thompson, 1975)

When the resting neuron potential becomes sufficiently less negative, the cellular membrane depolarizes and the nerve cell may be said to fire. This nerve impulse consists of a rapid exchange of positive and negative ions across the neuron membrane in order that the resting potential may be once again restored. Once initiated, the nerve impulse is conducted in both directions relative to the center of the neuron cell. However, the nerve impulse is transmitted beyond the membrane in only one direction, from the axon of the firing cell to the cell body or fibers of another cell. The amplitude and the speed of conduction of the nerve impulse is proportional to the diameter of the axon in which the impulses are generated. Because the extended axon tends to narrow and branch as it leaves the cell body of origin, the amplitude and the speed of the propagated impulse begins to diminish. Therefore, chemical mediators are needed at the terminals of the axon fibers to amplify the original electrical signal so that it can be communicated across the structural junction between the neurons. (Schwartz, 1973)
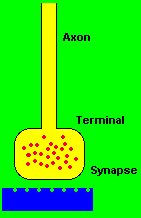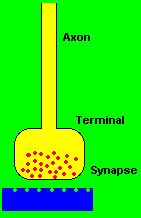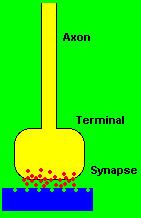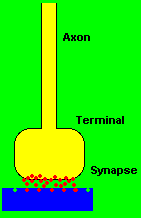
The junctional site of electrochemical transmission from one neuron cell to another neuron cell is called the synapse. The synapse is not an actual connection between cells, but rather a close approximation to one. The receiving end of the synaptic junction is usually located on the soma or dendrite fibers of neuron cells, whereas the transmitting portion is formed from the endings of small terminal fibers of the axon. The minute space between the two almost adjoining surfaces measures at about twenty millionths of a millimeter. The transmission of an impulse signal happens at the presynaptic terminal endings of the axon by the ejection of a chemical substance called neurotransmitter into the gap or junction between the two nerve cells. The neurotransmitter diffuses rapidly across the synaptic junction to the other side which is sensitive to the neurotransmitter and responds by initiating an electrical change at the postsynaptic region. The multiple branching of presynaptic axon terminals toward the multiple postsynaptic dendrite arborizations creates the neuron chain networks that constitute the electrochemical circuitry of the nervous system. (Corning & Balaban, 1968; Thompson, 1975)
The nerve impulse or propagated action potential operates according to an all-or-none spike discharge. This means that a neuron cell responds with the total voltage change that its polarized condition permits, if and only if, the electrical changes initiated at its membranous periphery reach a firing intensity or critical threshold value. The all-or-none spike discharge reflects the digital computation mode of the neuron cell. Like the elemental data in a digital computer that can be in only one of two possible states (yes or no), the elemental neuron can only be in one of two possible conditions (resting or firing). Once a spike, discharge is initiated, the impulse is propagated along the axon fiber by ion shifts in the Cell membrane down the full length of the axon. The nerve impulse normally lasts almost exactly the same length of time at each point in its journey along the axon fiber. Thus, repolarization of the axon fiber begins to spontaneously occur at the point closest to the main Cell body and then spreads progressively along the axon, moving in the same direction as the spreading depolarization. Guyton, 1972; Thompson, 1975)
The decision made by a neuron to fire or not to fire depends on those processes associated with the continuous electrical shifts induced on the cell membrane via the multitude of synaptic terminals impinging on its soma and dendrite fibers. The continuous intensity of these electric shifts is proportional to the amount and kind of incoming synaptic transmissions. Some neurons may have hundreds or even thousands of synaptic terminals impinging on them from other nerve Cells which are constantly ejecting various kinds of neurotransmitter. This kind of' multitudinous and continuous synaptic activity is known as the graded membrane potential of the neuron cell. Each neuron has the unique capacity to integrate all the incoming synaptic transmissions and establishes an overall electrical average which can then be expressed in terms of membrane potential level. If the average sum of the electrical shifts reaches the critical threshold value, the resting state is disrupted and a spike discharge occurs. On the other hand, if the average sum of the electrical shifts does not reach the necessary intensity, the resting state is maintained and a spike discharge does not occur. Therefore, like the analog computer that processes data that varies continuously, the neuron cell has the capacity to integrate the multitude of synaptic transmissions on a continuous basis before making the crucial decision to fire or not to fire. (Primbram, 1971; Schwartz, 1973)
The communication system between neuron cells, therefore, utilizes a signaling process that may be understood in terms of an analog-digital sequence. The analog process, as we have seen, involves the continuous chemical production and ejection at the synaptic terminals impinging upon the nerve cell. The digital process, as we have also seen, involves the discrete spike discharge that is characteristic of the nerve impulse. This kind of binary neural process also takes place at the peripheral and deep receptor sites where physical energies become transformed into electrochemical activity. The modified nerve fibers and specialized cells which are known as receptors evidence a high degree of sensitivity to certain forms of energy that they have been individually evolved to deal with. In the case of receptor cells, activation produces graded electrochemical activity that may be called the receptor potential. The activation of this graded receptor potential in turn activates a neurotransmitter across the micro juncture between the presynaptic receptor region and the postsynaptic dendrite fibers of the sensory neuron. In the case of modified nerve fibers, activation produces graded electrochemical activity that is known as the generator potential because the nerve fibers themselves generate the voltage change in the nerve cell body of origin. A major point to keep in mind is that in receptor systems there are receptor cells yielding receptor potentials and modified nerve fibers yielding generator potentials. However, both potentials may be subsumed under the general heading of graded electrochemical activity, which, if sufficiently large enough, will reach the firing threshold and initiate an all-or-none spike discharge in a sensory neuron. (McGaugh, 1971; Thompson, 1975)
Now in terms of influencing other neurons, it should be clear that a neuron can only choose to increase or decrease the probability of actualizing a spike discharge within itself or another nerve cell. Consequently, all the electrochemical effects at the synaptic junctions can be described in terms of two processes, excitation and inhibition. The biochemical mechanism by which these processes take place is through the presynaptic secretion of certain substances called the excitatory and inhibitory neurotransmitters. When the spike discharge spreads over a presynaptic terminal, the membrane depolarization initiates the ejection of one kind of chemical substance or the other into the synaptic space. If an excitatory neurotransmitter is released at the micro juncture, an immediate increase in the membrane permeability of the postsynaptic region occurs. This allows positive ions to flow more freely from the external biochemical fluid to the intracellular environment, which in turn causes the decrease of the electronegative charge inside of the cell's postsynaptic region. The decreased electro- negativity spreads throughout the nerve cell body because of the high conductive potential of the intracellular environment. Therefore, the actual effect does not occur only in the immediate vicinity of the synaptic juncture.
When the sum of the excitatory effects upon a neuron become sufficiently potent to initiate a change of about ten millivolts to the threshold level of minus sixty millivolts, a spike discharge takes place. An inhibitory neurotransmitter operates according to the same principle as an excitatory neurotransmitter except that the former increases the negativity of the intracellular environment by decreasing the degree of membrane permeability to positive ions beyond the resting state. A major point to keep in mind is that both inhibition and excitation usually work in conjunction with each other. For example, if the sum of inhibitory effects on a particular neuron shifts the electronegative charge within the cell towards the opposite direction of minus seventy five millivolts, then a change of plus ten millivolts brought about by the sum of the excitatory effects on the same cell would not be sufficient to reach the spike discharge threshold. In this way, neurotransmitters help to regulate neuronal activity, inhibiting certain neurons and exciting others, so that the central and peripheral nervous circuits stop and start as they should. (Guyton, 1972; Thompson, 1975)
Constant Spread of the Biogenetic DNA Potential
Each one of the twenty-six billion neuron cells that comprise the human nervous system is made up of a microscopic mass of living protoplasm bounded by an elastic plasma membrane. At the center of every nerve cell is a complex nucleus which is itself enclosed in a definite membranous tissue that separates it from the rest of the cytoplasm. Within the nerve cell nucleus is the master genetic material of the human psychobiology. This genetic material, which is also found in the other cells of the body, is composed of a formidable biochemical substance called deoxyribonucleic acid or DNA for short. The nerve cell nucleus also contains another kind of biochemical substance known as ribonucleic acid or RNA for short. DNA actually directs the internal life of every neuron cell manufacturing the biochemical messenger RNA that passes from the nucleus into the surrounding cytoplasm where it can then affect the synthesis of proteins, the physical and chemical changes in the pre-synaptic terminals, and the chemical reactions by which energy is provided for the vital metabolic processes.(Gaiton, 1966; Noback, 1975)
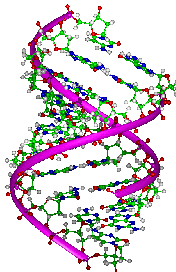
The DNA substance is fundamentally a long and thin chain-like molecule that is composed of a large number of smaller units called nucleotides. The nucleotides constitute the basic structural links in the complex DNA molecule. There are actually two polynucleotide chains in the DNA molecule that are wound together around an empty cylindrical space. Hence, the DNA molecule is not merely two parallel chains of matter but a highly organized structure shaped like two coiled wires in an electrical flex. This double helical molecule is enclosed in a definite protein coating specifically designed to fold and protect its polynucleotide chains that are many times as long as the soma or nerve cell body in which they appear. In fact the two polynucleotide chains in a single neuron cell would reach, nearly six feet if stretched full length. This means that the human nervous system alone contains approximately 29,545,455 miles of deoxyribonucleic acid. (DuPraw, 1970; Moore, 1962)
Based on my own meditative and intuitive understanding of the DNA molecule, I hypothesize that the interconversion of electrical and chemical energies deep within the coiled polynucleotide chains propagates a spinning electrical current which is the sine qua non of all living matter and that such a current precedes the evolutionary emergence of all forms of cellular life. This dynamic spinning life current fills the empty cylindrical space of the DNA molecule and transforms its coiled structure into a double helical heart. The image of the heart is appropriate here because the spinning DNA current is a complementary manifestation of a continuous electrical wave front and a discontinuous electrical pulsation that surges in two opposite directions one from the biogenetic dynamo to the cellular periphery and the other from the cellular periphery to the biogenetic dynamo. As the life current oscillates between the DNA molecule and the peripheral plasma membrane, it transforms the totality of the cellular protoplasm into a charged undulating substance. Hence, I am distinguishing between two kinds of pulsatory undulations in each living neuron cell, the rhythmical wavy thrusts of the DNA current and its effect the charged undulations of the protoplasmic body fluid. (De Ropp, 1972; Reich, 1970 & 1973)
Here is an Animated Image to Help
You Visualize the DNA Current

Animated gif by
mk-energies.com
The constant arrival and departure of the wavy throbbing life current from the DNA heart of every neuron Cell constitutes the biogenetic potential of nervous system functioning. This biogenetic potential is not just a question of the absence or presence of an electric intra-neuronal signal, nor is it a question of the amount of electrical shifts across the intracellular membrane. The significant electrical event here is the constant spread of a biogenetic current potential throughout the full extent of the internal cellular environment. The effectual charge intensity of the DNA current upon the cellular protoplasm is directly proportional to the amount and kind of electrical charges, positive and negative to neutral equilibrium, which are continuously propagated deep within the double helical dynamo. The effectual charge intensity is in a perpetual state of indeterminacy precisely because the random interconversion of electrical and chemical energies within the polynucleotide coils is specifically oriented toward avoiding a homeostatic state of charge equilibrium. This ever present recalcitrance to energy charge stabilization establishes a varying electrical activity that may be called the variable dynamo potential, for the double helical dynamo itself generates the perpetual voltage change in the undulating protoplasm of every neuron cell. What this all means is that the pulsatory DNA current manifests an increasing tendency to emphasize its positive or negative charge capacity as it streams through the cellular protoplasm developing an intra-neuronal phenomenon that resembles a prepotent charge wave front. At one moment, the pulsating wave front of the biogenetic current may emphasize a positive electrical charge, while at another moment it may emphasize a negative electrical charge. (Becker, Reichmanis, Marino, & Spadaro, 1976)
The constant spread of the variable dynamo potential serves to influence neuron cell firing by altering the resting electronegative charge of the intracellular environment. This means that the DNA current can increase or decrease the probability of actualizing the phenomenon of spike discharge by emphasizing its positive or negative charge capacity. Consequently, the intracellular effect that the DNA current has on neuron cell functioning may be described in terms of biogenetic excitation and biogenetic inhibition. When the DNA current emphasizes its positive charge capacity with a sufficient potency to initiate a change of about ten millivolts in the intracellular environment to the threshold level of minus sixty millivolts, a spike discharge occurs. However, when the DNA current emphasizes its negative charge capacity with a sufficient potency to initiate an increase in the electro negativity of the intracellular environment, a spike discharge will not occur. This means that the peripheral neuron membrane will respond to the DNA current with a total voltage change that its ionically polarized condition permits, if and only if, the predominate charge capacity within the life current becomes sufficiently more positive. (Pribram, 1971)
What I have described thus far serves to draw a distinction between a primarily external activation of neuronal impulses and a primarily internal activation of neuronal impulses -- a distinction which points to the fullest potential range of objective stimulus response and the fullest potential range of subjective stimulus response in human nervous system functioning. A major point to keep in mind, however, is that the external and internal factors that influence spike discharges along the various neuronal pathways work in conjunction with each other. For example, if the variable dynamo potential becomes sufficiently more negative to alter the electronegative charge in the intra-neuronal environment to minus seventy five millivolts, then a change of plus ten millivolts brought about by the sum of the external graded effects on the same cell body would not be sufficient to reach the spike discharge threshold of minus sixty millivolts. On the other hand, if the variable dynamo potential becomes sufficiently more positive to alter the electronegative, charge in the intra-neuronal environment to minus sixty five millivolts, then a change of plus five millivolts brought about by the external graded effects would be sufficient to reach the spike discharge threshold and cause the nerve cell to fire. What all this means is that the variable dynamo potential has the inherent capacity to regulate spike discharge activity by inhibiting certain nerve cells and exciting others throughout the full extent of the human nervous system. (Noback, 1975)
Hidden Variable of Quantum Transmission Potential
From what we know thus far in molecular psychobiology it seems reasonable to assume that all that we are as living organisms and our relation to all that has gone before us along the terrestrial evolutionary trajectory lies in each one of the double helical dynamos. More specifically, the DNA molecule in every neuron cell contains within itself the total informational residue of the evolutionary life process -- an immense coiled reservoir of latent engramic traces which includes not only the structural and experiential code of any given human organism, but also the accumulated informational input of the species as it evolved from the most simplest life forms (Samuel, 1972) . In this respect the variable dynamo potential of nervous system functioning also represents the potential emergence of informational impulses associated with the amoeba in us, the plant in us, the insect in us, the reptile in us, the fish in us, the fowl in us, the mammal in us, and the human in us. (Kendrew, 1966; Lilly, 1972)
Biogenetic Evolutionary
DNA Informational Potential

The limitless diversity of life, its orderliness, its forms and continuity is coded, controlled, and conveyed by those infinitesimal specks of material coil deep within the nucleus of every neuron cell body. This is to say that the two tiny spirals of matter bear within themselves the design and plan which accounts for the complexity of all that is living on our planet (Moore, 1962). Hence, the DNA molecule is not just a physical structure per se, for its enormous evolutionary informational potential gives it an additional meaning in terms of vast creativity, informational processing, and memory. I postulate that this extended meaning of the double helical structure is precisely possible because it employs a primitive storage, coding, and transmission system which antedates and is basic to all life forms from the simplest to the most complex. The storage, coding, and transmission system I am making reference to here involves the most fundamental level of mass-energy functioning which is responsible for the structural formation of the double helical dynamo and the actual manifestation of its throbbing electrical current. This deepest level of mass-energy functioning within the DNA molecule and its spinning pulsatory life current is made up of an immeasurable number of subatomic energy units which physicists usually refer to as quanta.(Bergstein, 1972; McConnell, 1968)
Based upon my own meditative and intuitive understanding of dynamic quantum functioning, I hypothesize that quanta can be conceptualized as subatomic bundles of swirling or spinning vortical energy systems that have a space aspect and a time aspect. Space expresses the extension of quanta within discrete and relative boundaries of swirling energies, while time expresses their variable duration as well as their causal interaction. In other words, quanta are energetic entities that are generally in one place at one time and cannot share the same place at the same time within the double helical dynamo. These vortical bundles of swirling energy unite to form the stable nuclear and atomic forces that actually build up the DNA molecule and give it its microscopic quality of helical structure. (Capra, 1975). However, within its helical structure, quanta function as independent wholes of swirling and spinning paradoxical activity each of which reflect a complementary manifestation of a vibratory energetic frequency and an undulatory energetic oscillation (Hoffman, 1959). The undulatory oscillation or wave nature of a quantum of energy varies with the velocity of its vortical spin. The greater the velocity of the vortical spin the shorter the wavelength, while the lesser the velocity of the vortical spin the longer the wavelength. On the other hand, the vibratory frequency of a quantum of energy varies with the mass-density of its spining vortical form. The higher the frequency of energetic vibration the more negligible the quantal mass-density, while the lower the vibratory frequency the greater the mass-density of the spinning energy quantum. (Kuznetsov & Yonovskii, 1965)
My current view of quantum functioning within the DNA molecule leads me further to hypothesize that innumerable bits of evolutionary information are coded through the complementary interplay between the two very different forms of energetic quantum action. What I am saying here is that the frequency of vibration in relation to the undulatory oscillation is equivalent to an energetic informational field of paradoxical action. The fact that a quantum of energy is equivalent to a certain amount of evolutionary information means that it can no longer be seen as just a bundle of spinning energy. It has to be also conceived of as a basic codon of the enormous data potential within the DNA molecule. The informational content of any given quantum therefore would refer to the particular manifestation of a spinning vibratory-wave combination out of all possible combinations which may potentially occur. Looked at in this way, a quantum of energy cannot be separated from its spinning informational field of paradoxical action. And likewise, the informational field of spinning paradoxical action cannot be separated from the distinct space and time in which the quantum of energy occurs. In this respect we can understand space, time, energy, motion, and information as inseparable and interdependent aspects of the most fundamental level of quantum functioning within the DNA molecule and its throbbing life current. (Capra, 1975)
The spinning informational fields of paradoxical quantum action which comprise the double helical dynamo and its pulsatory electrical current are extremely susceptible to influence from one another and to influences precipitated in the extra-cellular environment. I postulate that this interactive susceptibility is the essence of information transmission between the DNA molecule, its throbbing electric current, and the external synaptic terminal sites of every neuron cell body. The actual transmission of information can be understood as occurring through the mutual modulation of the interacting spinning quantal fields of paradoxical energetic action (Pribram, 1971). More specifically, the DNA helical configuration of quantum energy informational field patterns is continually supplied with incoming sensory data through quantal modulations in the pulsating electrical current which are themselves induced by quantal energy shifts at the peripheral membrane of the neuron cell. Likewise, the exterior synaptic terminal sites of every neuron cell body is supplied with emergent biogenetic data through quantal modulations initiated by the DNA molecule and its pulsating current. This means that it would be reasonable to assume that the quantal transmission process takes place at the peripheral and deep receptor sites of the sensory system where outside quantal codons become transformed into the graded neuronal activity which is itself comprised of vortical informational fields of spinning paradoxical energetic action. That graded electrical activity initiated at the deep and peripheral receptor sites yields an external quantum transmission potential, while the varying electrical current activity propagated by the DNA molecule yields an internal quantum transmission potential. However, both potentials may be subsumed under the general heading of "quantal transmission potential". (Gaiton, 1966; Guyton, 1972)
The quantal transmission potential necessarily implies that the dissemination of information throughout some twenty six billion neuron cells which build up the human nervous system as a whole is fundamentally a discontinuous process characterized by a finite number of quantal modulatory effects. It seems reasonable to assume however that there is a crucial factor operative at the quantal level of nervous system functioning which establishes the continuity, wholeness, and integration of that massive discontinuous information process (Sperry, 1970). I postulate that the operation of such an integrative informational factor is exactly possible because of a variable hidden within the apex of all spinning vortical quanta including those which effect the human nervous system from within and from without. Physicists understand this hidden variable I am making reference to here as a vacuum or void of potentiality from which all spinning vortical bundles of energy emerge and into which they seemingly vanish. They assume that there is literally no-mass in the vacuous apex of energy quanta that can 'be measured, no-motion that is commensurate with any unit or division of time. This does not mean that physicists view the vacuum as a state of mere nothingness, for it is believed to contain all energy quanta that exist in the present, have existed in the past, or will exist in the future (LeShan, 1974). They posit that the total energy-information of an isolated quantum is always conserved by the vacuum irrespective of whatever changes that may be initiated within it regarding the disappearance of the isolated quantum in one subatomic form and its reappearance in another (Gerholm, 1967). Going one step further here, I postulate that the ceaseless creation and absorption of quantal codons by the vacuum does not negate any bit of information involved at the deepest level of nervous system functioning in the sense of its becoming lost, gone, or non-existent. Such is necessarily the case since I have already equated energy with information. Hence, the conservation of information like the conservation of energy is conceptualized here as being fundamental to the regenerative potency of the vacuum found within the apex of every spinning quantal codon.
The hidden quantum variable or "vaccum of potentiality" belongs to a categorical referent which is beyond our present physical conceptions of space, time, motion, causality, and energy (Wigner, 1970). However, this does not mean that the hidden quantum variable is supernatural, but rather that it is impossible to detect with any current technological tools of experimental observation. In other words, we have not found a way as yet to objectively distinguish a hidden quantal phenomenon that is simultaneously operative in the human nervous system as a whole, the instruments of observation, the performance of those instruments, and the conditions of the experimental situation itself. What all this means is that the hidden quantum variable cannot be understood within the objective methodology of physical science which necessarily bifurcates the knower from the phenomenon to be known (Walker, 1974). Faced with an operative reality factor involved in nervous system functioning which lies beyond our extensional sensory observation, I adopt the intuitive unified cognition of meditation which fuses the knower with the phenomenon to be known in order to leap across our objective methodological limitation that presently restricts us from developing a preliminary hypothetical description of the aforementioned variable hidden in the apex of all spinning quantum vortical codons.

I perceive the hidden variable to be common to and operative in all those quantal codons which effect the deep and peripheral receptor sites of the sensory system as well as those quantal codons which comprise the DNA molecule and its throbbing electrical current. I hypothesize that the hidden quantum variable is essentially an analog of vacuous pulsation that evidences an undulatory motion of boundless magnitude oscillating at infinite velocity in alternately opposite directions from the apex of every vortical quantum involved in nervous system functioning to all other quantal codons in existence. The undulatory motion of the vacuous pulsation oscillating at infinite velocity diffuses the informational field of every quantal codon which participates in intra-neuronal and extra-neuronal potentials over illimitable distances in space and across past, present, and future aeons of time simultaneously while still conserving their local informational fields of paradoxical action (Taff, 1975). Our logical and analytical functions naturally find this simultaneity of informational diffusion and informational localization most difficult to comprehend. But perhaps we can clarify this enigma by imagining the hidden quantum variable as a pulsating vacuous point of infinite informational potentiality operative everywhere at the deepest quantal level of nervous system functioning. What is informationally potential, as I employ the concept here, may or may not become actualized at some point of time and in some field of space within the nervous system as a whole. This means that the hidden quantum variable constantly operative during all quantal transmissions introduces informational ubiquity, originality, freshness, randomness, and strangeness as ever present experiential potentialities emergent from the deepest reality level of human nervous system functioning. (Lilly, 1972; Gowan, 1974)

Art by Daniel B. Holeman
awakenvisions.com
The hidden quantum variable as I have described it here involves two simultaneous meanings of one and the same subatomic quantum reality process. In one meaning it represents an infinitely expanding informational continuum, while in another meaning it represents the vacuous integrating factor at the deepest quantal level of the human nervous system. What this all means is that the totality of the subatomic informational level of human nervous system functioning flows together in a continual rhythmic unison with all other quantal codons present throughout all space and time via the integral and vacuous pulse at the vortical center of every spinning quantum codon. That same vaccuous pulse may be likened to a throbbing cosmological heart that embodies the principles of omni-expansion and omni-contraction. According to this view, therefore, the informational continuity, wholeness, and integration of all quantal transmissions happening at any given moment in human nervous system functioning is not dependent on the complexity of cerebral organization, but is rather determined by the One rhythmical vaccous pulsation that is the omnipresent heart of an information gathering, processing, and endlessly evolving Cosmos (Lilly, 1975).
Summary of Major Points
The first section of this article revealed that the human nervous system can be divided into these two major categories: 1) the central nervous system which is comprised of the brain and the spinal cord; and 2) the peripheral nervous system which is composed of chains of nerve cells leaving the brain and the spinal cord. The peripheral system was subdivided into the somatic nervous system and the autonomic nervous system. I mentioned that the former is responsible for sensory reception and bodily action, while the latter serves to innervate the internal organs, the involuntary musculature, and the secretions of many glands. I showed how the autonomic system itself was further subdivided into the sympathetic and parasympathetic nervous systems which generally exert an opposing influence on the same organs, muscles, and glands. I depicted the somatic system as a complex nervous structure that is made up of afferent sensory pathways and efferent motor pathways. I said that the former transmit excitations from the ,sensory receptor mechanisms deep within the anatomy and at the body periphery to the central nervous system, while the latter transmit excitations from the central nervous system to the appropriate glands and muscles.
The second section of this article revealed that the fundamental functional units of the human nervous system are the neuron cells whose semipermeable membranes are in immediate contact with a surrounding biochemical fluid composed of electrically charged ion molecules. I pointed out that the porous nature of each one of these nervous cellular membranes allows negative ions to pass freely while restricting the passage of most positive ions. The attendant electrical polarization between the more negative intracellular environments and the more positive extracellular fluid creates resting electrical potentials across the surface of the neuron membranes. I showed how resting neuron cells will necessarily respond with impulse firings or propagated action potentials when their intra-cellular environments become sufficiently less negative to reach a critical threshold value. The impulse firing is essentially a rapid exchange of positive and negative ions across the neuron membranes in order that the resting potentials are once again restored. I mentioned that the information contained in the impulse firings is passed on between neuron cells via the ejection of electrochemical substances at the minute synaptic regions where they almost come into immediate contact with one another. I described how physical energies impinging on the deep and peripheral receptor sites become transformed into an electrochemical informational activity at the synaptic terminal sites of afferent sensory neurons leading to the spinal cord and brain.
The third section of this article postulated that the constant arrival and departure of the throbbing life current from the DNA heart of every neuron cell constitutes the biogenetic potential of nervous system functioning. The effectual charge intensity of the spinning biogenetic current upon any intracellular neuron environment is directly proportional to the amount and kind of electrical charges, positive and negative to neutral equilibrium, that are continuously propagated deep within the coiled polynucleotide chains of the DNA molecule. I stated that the pulsatory DNA current of each neuron cell manifests an unceasing tendency to emphasize its positive or negative charge capacity as it streams throughout the cellular protoplasm developing an intracellular phenomenon that resembles a prepotent charge wave front. It was pointed out that the constant spread of this variable biogenetic current potential serves to influence neuron impulse firing by altering the resting electronegative charge of the intracellular environment. More specifically, I emphasized that the throbbing DNA current can increase or decrease the probability of actualizing the propagated action potential of every neuron cell in the human nervous system by emphasizing its positive or negative charge capacity. I then concluded by positing that the internal and external factors which influence the propagated action potentials throughout the full extent of neurological functioning work in conjunction with each other to create the fullest personal range of objective sensory experience and the fullest evolutionary range of subjective biogenetic experience.
The fourth and final section of this article indicated that the enormous evolutionary informational potential inherent in the DNA molecule gives it an additional meaning in terms of an infinite creativity, processing, and memory. I hypothesized that such an extended meaning was precisely possible because the double helical dynamo employs a primitive storage, coding, and transmission system which involves its most fundamental level of quantum mass-energy functioning. I stated that the bits of evolutionary information are coded through the complementary interplay between two very different forms of energetic quantum action. More specifically, I posited that the frequency of quantal vibration in relation to the undulatory quantal oscillation is equivalent to an energetic informational field of paradoxical action. The informational content of any given quantum of spinning paradoxical action was associated with any particular manifestation of a vibratory-wave combination out of all possible combinations which may potentially occur. I postulated that the information transmission between the DNA molecule, its throbbing electrical current, and the external synaptic terminal sites of every neuron cell occurs through the mutual modulation of the interactive quantal fields of paradoxical energetic action, it was pointed out that the DNA molecule as an energetic helical configuration of quantum information field patterns is continually supplied with incoming sensory data through quantal modulations in the pulsating electrical current which are themselves induced by quantal energy shifts at the peripheral membranes of all neuron cells. I stated that the essence of this quantal transmission potential necessarily implies that the dissemination of information throughout the full extent of the human nervous system is fundamentally a discontinuous process characterized by a finite number of quantal modulatory effects. However, I went on to hypothesize that there is a hidden variable hidden in the spinning apex of all vortical quanta in existence which establishes the continuity, wholeness, and integration of that massive discontinuous information process. I revealed that the hidden quantum variable is essentially an analog of vacuous pulsation that evidences an undulatory motion of boundless magnitude oscillating at infinite velocity in alternately opposite directions from the apex of every vortical quantum involved in nervous system functioning to all other quantum informational codons in existence. The hidden quantum variable was identified as the operative reality factor that diffuses the informational field of every quantal codon which participates in intra-neuronal and extra-neuronal potentials over illimitable distances in space and across past, present, and future aeons of time simultaneously while still conserving their local informational fields of paradoxical energetic action. I went on to emphasize that the vacuous pulsatory variable hidden in the spinning apex of all vortical quanta in existence functions as an infinitely expanding informational continuum as well as an operative integrating factor at the deepest quantal level of human nervous system functioning. I concluded by hypothesizing that the hidden quantum variable establishes the informational wisdom, integrity, and wholeness of the human psychobiology and individual personality before the evolution of the cerebral cortex and its emergent processing function known as thinking mind.
International Copyrights
Dr. Bonnici. January 28, 1978/2020
|